https://zhuanlan.zhihu.com/p/461143839
题记
根据接触很多孩子的经验,而不是根据理论,我们认为这个男孩的正面和负面特征是天生的、必然的,它们连接而形成了真正的平衡人格中两个必要的部分。我们也可以这么形容,这个男孩面对自己以及外在世界所遇到的困难,是因为他拥有上天赐予的特殊天赋而必须付出的代价。
—— Hans Asperger, 1938
*封面图片 Groc vermell blau - Vassily Kandinsky, 1925
0 绪论
0.1 说明
版本说明:2023年第一次修订版。主要改动:新增1.6节,新增2.1节,修正并合并5.1, 5.2节,新增5.2~5.6节,新增5.8节,新增第7章。
全文共81,679个字,阅读大约需要120分钟。
快速阅读路径(30分钟):选取以下章节1.1~1.3, 1.6, 2.1~2.5, 3.1~3.4, 4.1, 5.1~5.6
(跳过的章节用*表示)
极速阅读路径(15分钟):选取以下章节1.1~1.3, 1.6, 2.1~2.3, 3.1~3.4, 4.1, 5.1, 5.6
自我诊断(2个小时):第8章
目前国内中文的阿斯伯格相关文献过于稀少,主要的中文出版物与国际有着至少20年的差距,这导致了国内对于阿斯伯格群体的认识与发达国家相比有着非常大的差距。中国有着庞大的人口基数,以美国目前排查的自闭症谱系人口比例来看,中国大约有着3200万的谱系人口。如此庞大的一个群体在国内却没有与之体量匹配的重视程度。更不用说其间数不胜数的本该大大推进人类科技发展,却最终由于社会缺乏理解而被忽视湮没的高智力潜力(HIP)的阿斯伯格人士。因此我们亟需通过知识平台向公众普及阿斯伯格知识。
基于这个需求,本文从神经科学机理开始梳理一条脉络清晰地解读了阿斯伯格的表征,系统地呈现了自闭症谱系领域尤其是阿斯伯格领域近十年内的一系列重要进展,可视为最新版本的阿斯伯格完全指南。一些结果仍然是猜想,不过大多都是学术界普遍认同的。文章取得了一些高智力阿斯伯格的共鸣,但其对更广泛人群的适用性必须谨慎辨别。我本人是数学专业的,和神经科学或医学领域毫无交集,任何医疗方面的建议请务必找专业医生咨询确认。
为了文章更具有前瞻性,我们用越来越多的学者开始使用的ASC(自闭症谱系状况)一词来代替原来ASD(自闭症谱系障碍)一词,但在翻译过去的文献时,仍遵照原文的用词。
0.2 引言
在数十万年前,一群原始人中出现了一些全新的基因,使得人类第一次与尼安德特人等表亲产生了区分。其中一些关键的基因给人类带来了更高的智力,使人类成为了“智人”。与此同时,人类当中广泛出现了一种几乎不会在其他物种中出现的问题,被称为自闭症[1]。另一种假说是5到10万年前人类从非洲迁徙北上时与现已灭绝的更为聪明的尼安德特人发生了广泛的基因融合,获得了这些高智力基因,之后回迁的一部分人类又给非洲人类也带去了这种基因,这些基因导致了自闭症[2] [3] [4]。总之,从此以后,人类的文明突然以一种极为迅猛的速度发展,短短数万年就从到处跑的狩猎采集时代转变为自闭宅家种田的农业时代,再转变成人人抱着手机的信息时代,大大超过了之前百万年人类成就总和。一些人类开始宅在山洞成天刻板地磨着石器,这些人后来又整天一动不动天天观察着植物生长规律学会了种地,他们后来又开始痴迷于看星星,搞数学——甚至士兵攻到脚下也要解出一个几何题。他们开始观察着光影细微的变化画出了极为写实的油画,从无穷无尽的数字中发现了行星运行的模式,站在讲台上讲着微积分——即便台下一个人也没有,现在他们又成天窝在电脑前写着代码。他们一直在普通人身边,伴随着人类整个文明史。
我们现在知道,自闭症是一个连续谱系(自闭症谱系状况,ASC;或称自闭症谱系障碍,ASD),从对着黑板侃侃而谈的阿斯伯格到生活不能自理不会说话的典型自闭症,特质从轻到重,但最先引起人们关注的是一些几乎不会讲话的自闭患者。随后1944年,医生Asperger发现,那些几乎不会讲话的自闭症患者往往有着从事高智力工作的家属,自闭症开始和高智商基因联系在一起。他注意到一群古灵精怪的小孩和这些典型的自闭症患者有着诸多相似之处。他把他们描述为“缺乏同理心,几乎不会发展友谊,说话时不管对方感受,对某些东西强烈的兴趣,动作有些笨拙”。他注意到其中一个小孩成为了天文学家,解决了牛顿的一个错误。还有一个他的阿斯伯格患者Elfriede Jelinek获得了诺贝尔奖。同时人们观察到,一些传统上不被认为是阿斯伯格的普通天才儿童也出现了一些社会适应障碍,他们与阿斯伯格的实质上难以区分[5]。现在我们知道,自闭症等位基因和高智力等位基因广泛重叠[6],包括更大的大脑、快速的大脑生长、感觉和视觉空间能力的提高、突触功能的增强、注意力集中程度的提高、慎重的决策等等[5]。当高智商基因比较严重时,就会带来自闭症。特别严重的情况,高智商基因会带来一些严重的并发障碍,使人丧失语言[7]和社交功能。阿斯伯格就处于严重和不严重的边缘地带。
2019年Nature Neuroscience的文章[8]指出,神经干细胞阶段更加开放的染色质使得自闭症谱系人群(ASC)的神经细胞发育更庞大迅速。自闭症大脑各个部分以大大超过常人的规模发育,尤其是额叶[9]——这是大脑最高级的一些活动的生理基础,被认为是人类成为万物之灵长的关键。人类大脑皮层的前额叶部分大约是黑猩猩的3.6倍[10],而自闭症儿童前额叶皮层神经元数量达到了正常儿童的1.67倍[11]。半个物种级别的神经元规模和分叉密度差异使得自闭症感官和情绪异常敏感(敏锐),产生了与常人不同的感知模式[12],在全部的感官:视觉、听觉、味觉、嗅觉、触觉、前庭感普遍出现了极度敏感的现象。反应到临床上出现怕光、更强的视觉感知能力[13]、容易被惊扰、非常好的音感[14]、挑食、对衣服材质挑剔(以至于可能经常穿同样的衣服)等等现象。这种感知能力使得他们时常感到感官过载,和他人社交时被过量的社交情绪社交信息淹没。如同对光线敏感的人会跑到暗的地方一样,ASC们会跑到没有人的地方来避免信息过载。和人谈话时目光躲闪也是一样的原因,为了避免接受到过量的信息。为了简化感受到的过量信息,ASC们发明了“系统化”的办法[15] [16],即分析与构建系统的认知模式。他们一刻不停地寻找着信息的规律和模式,将各种东西分类,建立理论来预测周围的信息[17],防止被信息淹没。不喜欢环境变化、追寻秩序、刻板固执也是因为改变环境则会重新带来信息过载,需要花时间重新寻找模式,重新系统化。为了生存所被迫练出的系统化能力被运用在认识世界上,就成为了数学的公理化系统与科学研究的基本方法[18]。事实上,数学几乎就是完全由阿斯伯格建立起来的学科[19] [20] [21] [22],物理的那几个关键人物也和ASC跑不了关系[23]。剑桥大学教授Simon Baron-Cohen(主导了剑桥自闭症研究中心,现代众多ASC临床评估量表的作者)对剑桥数学系进行调查发现,剑桥大学本科生中数学系的阿斯伯格比例为其他专业的8倍[24],并且当时在剑桥的Fields奖得主Richard Borcherds就是典型的阿斯伯格[25]。
ASC大脑神经细胞的高度发育和人体其他生理结构的不匹配产生了一系列并发问题。例如位于脑干的蓝斑核主导的去甲肾上腺素神经调节系统(LC-NE)明显调节能力不足。ASC大脑有着远高于普通人群的神经元数量和突触间隙数量,但合成去甲肾上腺素神经递质的蓝斑核却与正常人无论是大小上密度上还是数量上都没有区别[26]。去甲肾上腺素能够增强对感觉输入的处理,增强注意力,增强长期记忆和工作记忆的形成和检索,并通过改变前额叶皮层和其他区域的活动模式,增强大脑对输入的反应能力。去甲肾上腺素转运的不足引发了ADHD/ADD(注意力缺陷多动障碍/注意力缺陷障碍)[27],这在许多阿斯伯格身上被观察到,以至于ADHD被认为可以作为ASC的连续变体[28]。LC-NE系统调节能力不足也导致了某些阿斯伯格情绪调节能力的问题、工作记忆问题和学习障碍[29]。由于情绪易感性和社会的不理解,1/3的阿斯伯格青少年和2/3的阿斯伯格会经历至少一次严重的抑郁。另外阿斯伯格神经递质浓度随年龄的变化可能带来焦虑[30] [31]。如何预防和解决这些问题是每个阿斯伯格必须学习的。
简单地说,ASC的诊断标志是:1.在开始和维持社会交流和互惠的社会互动方面持续存在缺陷。2.持续的限制性、重复性和不灵活的行为、兴趣或活动模式。ASC在发育期就会出现的,但特征可能要到后来社会需求超过其能力时才会完全表现出来。这些特征导致个人、家庭、社会、教育、职业或其他重要的功能领域出现重大障碍。一些人通过特别的努力,使得他们的缺陷对其他人来说可能并不明显,此时诊断仍然适用。具体的诊断需要一些量表、发育史等综合推断。阿斯伯格是ASC中语言与认知功能没有缺陷的一支。特别的语言和认知能力模式使阿斯伯格症有别于典型的自闭症:虽然阿斯伯格症患者的言语(对应知觉推理、言语理解)和操作(对应加工速度)智力都高于高功能自闭症(HFA),但言语理解智商得分明显较高,远高于HFA,甚至高于正常儿童平均水平一个标准差[32] [33]。这与阿斯伯格症儿童的刻板印象一致:像小教授一样使用非典型的学究式语言(话说回来,“教授”的刻板印象大概也是历史上的阿斯伯格教授们塑造的)。总之,阿斯伯格是一种高智力相关遗传特征的过度发育模式,结果没有使语言和认知功能低于正常人水平,但会产生社会交往困难和重复性行为或强烈固执的兴趣。
0.3 术语缩写字典
- 5-HT = 5-Hydroxytryptamine = 5-羟色胺 = 血清素
- AADC = Aromatic L-amino acid decarboxylase = 芳香族L-氨基酸脱羧酶
- ABA = Applied Behavior Analysis = 应用行为分析
- ADD = Attention Deficit Disorder = 注意力缺陷障碍
- ADHD = Attention Deficit Hyperactivity Disorder = 注意力缺陷多动障碍
- AQ = Autism Spectrum Quotient = 自闭症谱系商
- AS = Asperger Syndrome= 阿斯伯格综合征。 下文亦指代阿斯伯格人士。 |备注:在DSM-5诊断标准中阿斯伯格被归类为一种症状轻微的ASC。
- ASC = Autism Spectrum Condition = 自闭症谱系状况 | 备注:学者们对ASD的新式称法,基于最新的ASD不是“障碍”的潮流观点。
- ASD = Autism Spectrum Disorder = 自闭症谱系障碍 |备注:包含自闭症、亚斯伯格症候群、待分类的广泛性发展障碍。
- CBT = Cognitive Behavioral Therapy = 认知行为疗法
- CNS = Central Nervous System = 中枢神经系统
- DA = Dopamine = 多巴胺
- DBH = Dopamine β-Hydroxylase = 多巴胺β羟化酶
- DSM = Diagnostic and Statistical Manual of Mental Disorders = 精神疾病与诊断统计手册,由美国精神医学学会出版
- EEG = Electroencephalography = 脑电图
- EQ = Empathy Quotient = 同理心商数
- FQ = Friendship Quotient = 友情商数
- HFA = High-Functioning Autism = 高功能自闭症 |释义:一种自闭症分类,没有表现出智力障碍,但可能在沟通、情感识别和表达以及社会互动方面表现出缺陷。
- HIP = High Intellectual Potential = 高智力潜力 |释义:通常指韦氏智力大于或等于130的人所蕴含的智力潜力特质。
- HSP = Highly Sensitive Person = 高敏感人士
- LC = Locus Coeruleus = 蓝斑核
- L-DOPA = levodopa = 左旋多巴
- MRI = Magnetic Resonance Imaging =核磁共振成像
- NaSSA = Noradrenergic and Specific Serotonergic Antidepressants = 去甲肾上腺素和特异性5-羟色胺能抗抑郁药
- NDRI = Norepinephrine–Dopamine Reuptake Inhibitor = 去甲肾上腺素-多巴胺再吸收抑制剂
- NE = Norepinephrine = 去甲肾上腺素
- NT = Neurotypical = 神经典型发育 |释义:用来指非自闭症谱系人士,即神经结构与大多数相同的普通人,常脑人。
- PNMT = Phenylethanolamine N-methyltransferase = 苯乙醇胺N-甲基转移酶
- PNS = Peripheral Nervous System = 周围神经系统
- SNRI = Serotonin–Norepinephrine Reuptake Inhibitors = 5-羟色胺和去甲肾上腺素再摄取抑制剂
- SNS = Sympathetic Nervous System = 交感神经系统
- SQ = Systemizing Quotient = 系统化商数
- SSRI = Selective Serotonin Reuptake Inhibitor = 选择性5-羟色胺再摄取抑制剂
- WAIS = Wechsler Adult Intelligence Scale = 韦氏成人智力量表
- WISC = Wechsler Intelligence Scale for Children = 韦氏儿童智力量表
1 阿斯伯格的神经学机理
1.1 自闭症的核心是感知模式异常
C. Robertson和S. Baron-Cohen 2017年在Nature Reviews Neuroscience综述文章Sensory Perception in Autism[12]中指出,学者们越来越注意到自闭症的核心症状应该是非典型的感知模式,而社交异常只是感知敏感带来的附加产物。感知模式影响着生活的方方面面,与NT(常脑人)的感觉差异会导致与NT交流不在一个调上,最终导致社交隔离感。严重的情况甚至会引起语言隔离,导致典型自闭症。其感知模式也有更加关注细节的趋向。下面几段摘自论文:
反思我们自己和他人的想法和情绪的能力(即心智理论)是人类认知的一个决定性特征。患有自闭症谱系疾病的儿童在这种能力的发展上出现了延迟,对整个人生中的认知共鸣产生了连锁反应。有趣的是,这些社会认知的改变伴随着对世界非常不同的感知体验。据估计,多达90%的自闭症患者会出现非典型的感官体验,并影响到每一种感官模式:味觉、触觉、听觉、嗅觉和视觉。自闭症研究的一个核心挑战是确定将这些认知和感觉的不同方面联系起来的共同线索。哪些神经生物学的改变可能会影响到社会认知和感官知觉这样不同的过程?
最新的国际自闭症诊断标准强调了这一挑战,该标准现在将感觉敏感作为一个核心诊断特征。虽然在过去早期研究中的病情报告中提到了感觉症状,但它们历来被认为是自闭症认知的次要方面,而不是主要的表型标志。除了对创造自闭症友好环境有临床意义外,了解自闭症中感官差异的重要性,对自闭症的神经生物学描述至关重要。
然而,哪个才是首先出现的是关键。自闭症是否像人们常说的那样,是一种 "社会大脑 "的障碍,感觉上的差异是在人生中缺乏社交后的次要结果?或者它影响了社会处理和感觉处理的一般机制(如注意力)?或者感觉上的差异在发育和神经生物学方面都是主要的?
在此,我们探讨感觉特征是否是自闭症的核心表型标志。
最终,文章给出结论:
本评论认为,感觉症状是自闭症神经生物学的核心和主要特征。具体来说,自闭症的感觉处理差异在发育早期就很明显,最早可在婴儿期出现,并能预测儿童期后期的诊断状况。它们可以预测成人社会和认知功能的高阶缺陷,并解释诊断评估中社会和交流症状的独立差异。此外,自闭症相关的感觉症状反映了感觉专用神经回路的改变,包括大脑初级感觉区域的神经分子和解剖学变化,而不是高阶认知过程改变的次要后果。

1.2 更开放的神经干细胞染色质导致神经细胞发育更庞大复杂
AS在六种感官上表现出普遍的敏感,包括视觉、听觉、触觉、味觉、嗅觉、前庭觉。(某一些特别低敏理论上也会导致感知和他人不着调,但一般AS为高敏,某些个别感官的低敏也被认为是过敏导致的抑制(大脑会自发抑制隔离过于强烈的感受,属于保护机制),所以暂且讨论过于敏感的情况)这样普遍性的高敏感自然被认为是由大脑神经结构的普遍性差异导致。S.T. Schafer等人2019年在Nature Neuroscience一篇论文Pathological priming causes developmental gene network heterochronicity in autistic subject-derived neurons[8]中指出,(译自Science News)从ASC患者身上提取的早期发育阶段神经细胞比从没有ASC的人身上提取的细胞长得更大,分支更长、更复杂。由自闭症患者神经细胞衍生的三维有机体,也更加庞大。除了这种身体上的发展,一组对大脑发育很重要的基因也更早地开启了。
问题甚至在这些细胞即将成为神经细胞的时候就已经有了。在神经干细胞阶段,这些细胞的某些染色质比它们应该有的更开放和更容易接触,这种开放性可能导致基因表达异常活跃。加州大学戴维斯分校的神经科学家David Amaral说:
“结果显示,开放的染色质可以对神经元发育产生重大影响。”
在科学家们使之被迫跳过神经干细胞阶段时,细胞的异常生长模式消失了。论文合作者Rusty Gage说:
“对我来说,这个实验的成功令人惊讶。这一发现支持了这样一个观点,即在一个特定的时间发生的单独事件,即神经干细胞阶段的开放染色质,可能为大脑带来了自闭症。”
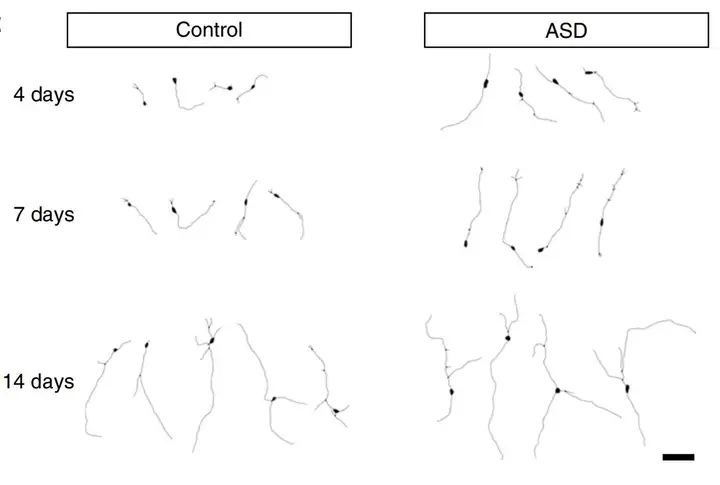
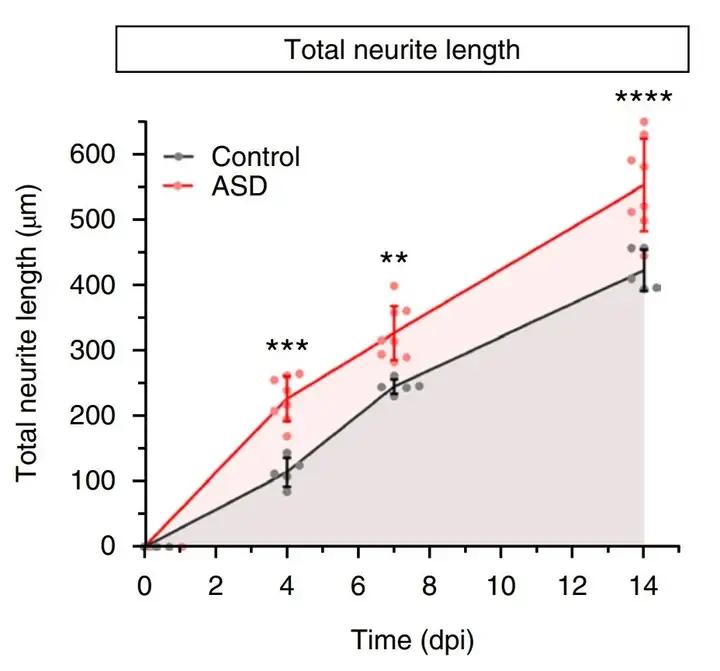
早期神经元阶段的异常基因连接发育动态与之前神经干细胞阶段的染色质可及性变化有关,跳过相关的神经干细胞阶段时,神经发育变得正常。
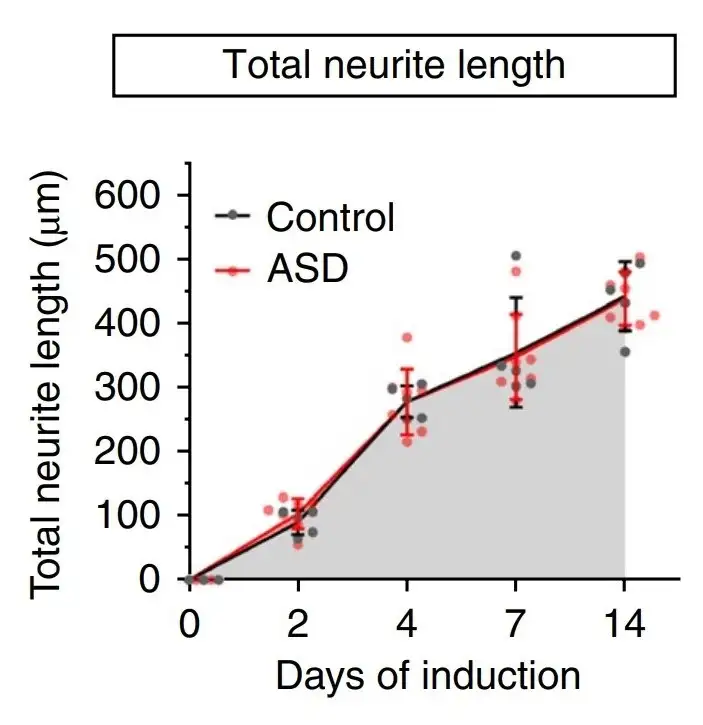
1.3 庞大复杂的神经细胞决定了阿斯伯格不同的感知模式
庞大复杂的神经细胞发育模式在大脑每个局部都形成了更多的联结,并且更多的联结使两个相邻神经元的联结更难完全消失。这解释了阿斯伯格以及自闭症谱系人群极强的机械记忆力与长期记忆力,也解释了为何同样的感觉信号在AS大脑内产生了更激烈的反应,使之感官更加敏感。
ASC人群有着更好的音感[14]:
患有自闭症谱系障碍的儿童在单音和旋律背景下表现出较高的音调辨别能力,以及对旋律的长期记忆能力。音调记忆与非语言流体推理能力的得分呈正相关。
更强的视觉感知能力[13]:
自闭症患者在进行视觉搜索、视觉辨别和嵌入式图形检测时,往往表现出更强的感知能力。同样,在执行一系列感知或认知任务时,自闭症患者比非自闭症患者表现出更强的视觉系统生理参与。
一种局部-整体连结数量假说被用来解释ASC的局部-整体感知模式。
单从数量上,虽然大多数实验数据表明ASC局部联结更多,而整体联结的数量在一些调查中少于NT,某些调查中多于NT,目前存在争议。甚至在局部-整体不同的定义和实验标准下有完全相反的结果(参考Dr. Caroline Robertson 2018年在Dartmouth的演讲14分13秒Autism: A view from neuroscience - A CCN public lecture)。但大体而言,目前学术界普遍的观点猜想局部连接的丰富性可能是自闭症成因的一种解释。这使得阿斯伯格等ASC人群有着更加关注“细节”的局部-整体感知模式。当整体联结相比局部联结稀少太多时,不同大脑区域间的联系活动相对于局部区域内大脑的活动显得更不活跃(简单地说,局部信息过于强烈淹没了整体信息),会导致感统失调。因此AS在肢体协调上通常更弱一些。更加严重时可能导致多感官结合的严重缺陷,比如使得听觉和视觉之间联系变得不那么紧密,这种视听之间的错位在视听语言范式中表现得特别明显,被认为可能是导致语言发展障碍的重要原因。(参考Stevenson, R. A.等人2017年的论文 The cascading influence of multisensory processing on speech perception in autism[7])在这种特别严重的情况下,患者出现与周围人的语言隔离,不再表现出阿斯伯格,而是表现出典型自闭症。
为了理解这种局部-整体的不同的感知模式,我们简单地建立一个模型不严谨地计算一下。同样大脑体积下,我们可以预测整体感知能力主要取决于大脑区域间单位截面积连接的神经元数量(大致正比于神经元总数量乘以单个神经元长度),而局部感知能力主要取决于相邻神经元间突触分支的密度(大致正比于神经元总数量乘以神经元三维有机体体积)(仅作理解用,暂无研究支持)。而目前的研究看来,AS在幼儿时期神经元数量比NT约高了65%,而成年时与NT数量上基本相同,但单个神经元分支数量上约为NT的2到3倍(参考Autism: An evolutionary perspective, Professor Simon Baron-Cohen, 1st Symposium of EPSIG, 2016 13分19秒)。非常不严谨地估算,当AS的神经细胞三维有机体的半径是NT的 倍(),总数量为NT的 倍()时,AS的局部感知力是NT的 倍,而整体感知力是NT的 倍。AS获得的局部信息和整体信息比是NT的 倍。这可以形象地解读为成年AS的局部感知和整体感知都大大强于NT,分别是NT的 和 倍,但局部感知比起整体感知强于NT更多,使得局部感知信息比整体的比起NT更为强烈,这种局部偏向性是NT的 倍。本质上,AS很可能只是被局部信息淹没了,整体的感知也比NT强。但这种注意力对局部细节的偏向性使得没有经过引导训练的AS把更多的精力投入到局部细节,而对整体不加以关注,久而久之对整体相关方面的累积的认识会更少。大脑由于其可塑性,在长期更多注意力放在局部活动的情况下,加强了局部联结,减弱了整体联结。这样的大脑偏向性训练最后也可能使得AS的整体感知能力反而不如NT。
1.4 自闭症神经发育的动态特征和大脑区域特征*
之前几节我们看到,近年的研究声明了干细胞阶段开放的染色质使得神经细胞发育更迅速,这种神经发育的动态特点早在2007年的综述文章Mapping Early Brain Development in Autism[9]中就在自闭症中被指出。该文章摘要写道:
虽然自闭症的神经生物学研究已有二十多年,但这些研究大多是在临床症状出现10年、20年或更长时间后才对大脑结构进行研究。导致自闭症的病理生物学仍然未知,但其特征可能在临床症状出现的头几年最为明显。这篇评论强调了关于生命最初几年的神经生物学发现,并强调早期大脑过度生长是自闭症病理生物学的一个关键因素。我们推测,神经元数量过多可能是早期大脑过度生长的一个原因,并产生了神经模式和布线的缺陷,局部和短距离的皮质相互作用旺盛,阻碍了大脑区域之间大规模、长距离的相互作用功能。
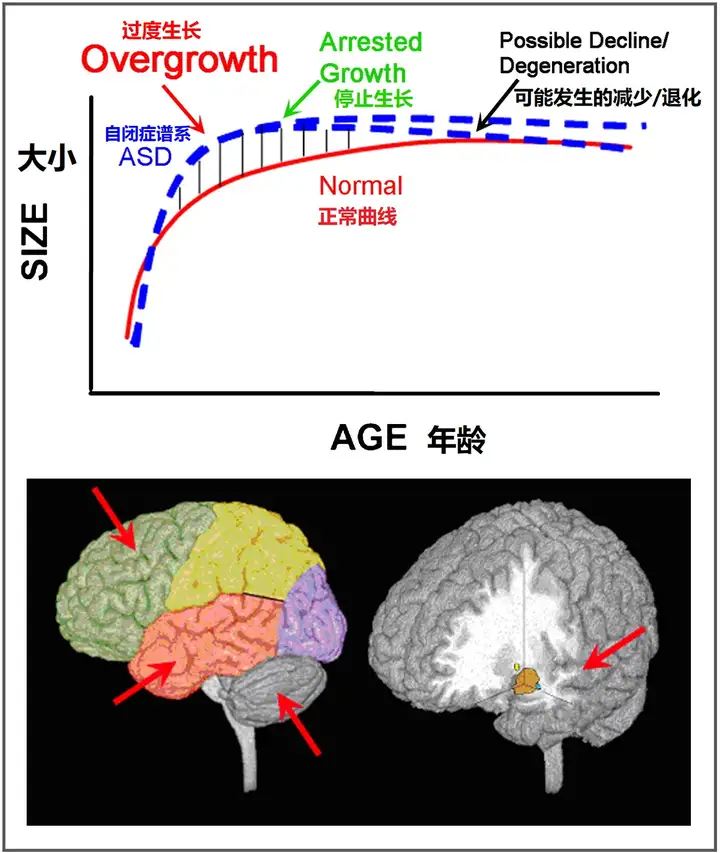
节标题:自闭症的早期大脑过度生长
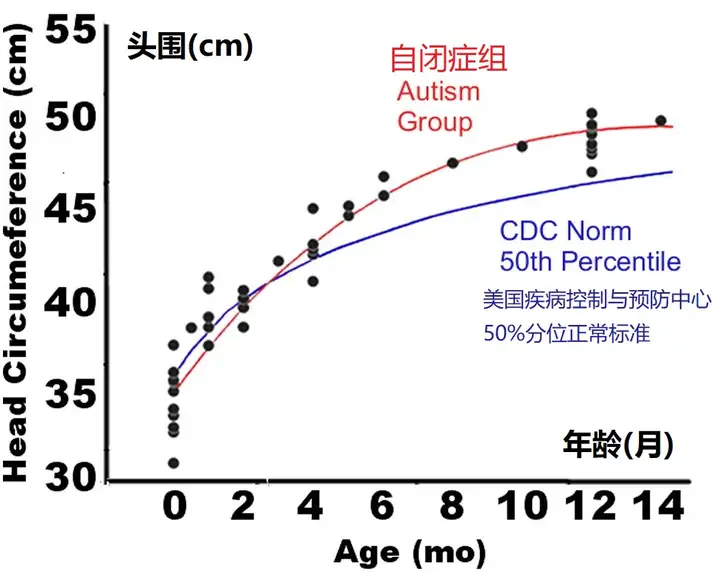
来自头围和核磁共振研究的证据
在生命的最初几年,头围与大脑大小有很好的相关性(Bartholomeusz et al., 2002),它已被用作自闭症相对大脑大小的回顾诊断指标。在出生时,后来发展为自闭症的婴儿的头围通常接近正常或略低于正常平均水平(Courchesne et al., 2003,Dawson et al., 2007,Dementieva et al., 2005,Dissanayake et al., 2006,Gillberg and de Souza, 2002,Hazlett et al., 2005,Lainhart et al., 1997,Mason-Brothers et al., 1990,Stevenson et al., 1997)。在提供个人数据的两项研究中(Courchesne et al., 2003,Dementieva et al., 2005),90%-95%后来被诊断为自闭症的新生儿在出生时有平均或略小于平均的头围,只有约5%在出生时有过大的头围。然而,Courchesne等人(2003)最近发现,自闭症患者到1或2岁时,头围(HC)会异常增大(图7)。这一早期过度生长的发现现在已被许多独立的研究小组所复制(Dementieva et al., 2005,Dissanayake et al., 2006,Hazlett et al., 2005,Dawson et al., 2007)。
当自闭症儿童长到2-4岁时,相对于正常发育的2-4岁儿童来说,整体的核磁共振脑容量异常增大了约10%(Carper et al., 2002,Courchesne et al., 2001,Hazlett et al., 2005,Sparks et al., 2002)。最近的两项核磁共振研究证实了年轻自闭症儿童大脑增大的发现,受试者的平均年龄为2.7岁(Hazlett et al., 2005)和3.9岁(Sparks et al., 2002)。对2005年初发表的所有儿童、青少年和成人的核磁共振脑容量数据进行的汇集分析表明,自闭症患者脑容量最大的时期是在幼儿期和儿童早期(但需要注意的是,即使在较大的年龄段,自闭症患者的脑容量仍有1%-3%的增长)。(图8)(Redcay and Courchesne, 2005)。
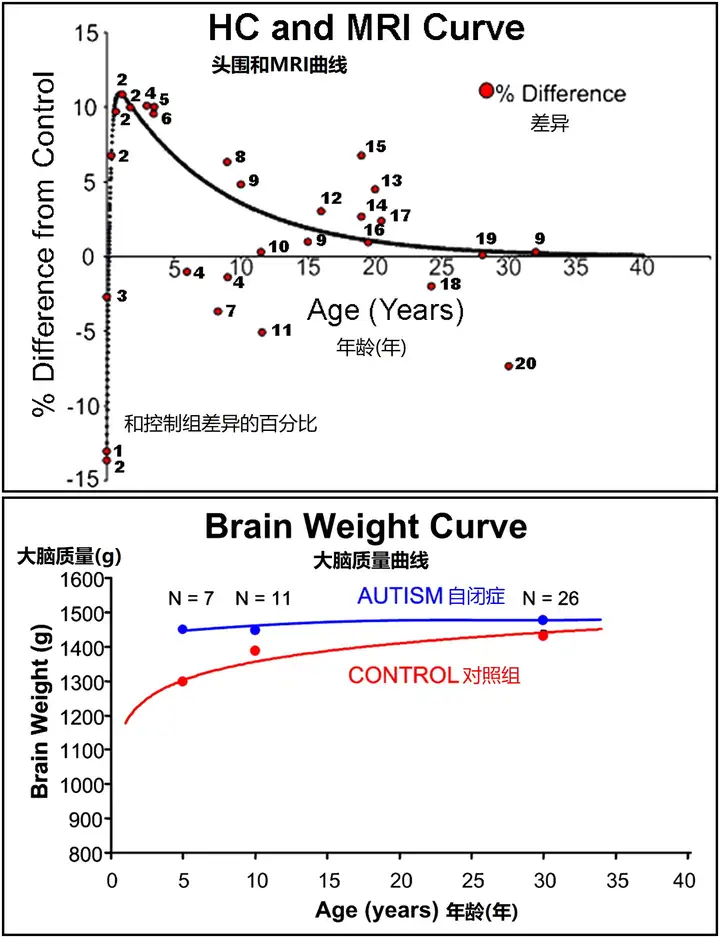
死后的大脑重量数据,虽然不一定总是可靠的措施,但确实证实了头围和MRI研究的结论。Redcay and Courchesne (2005)在唯一一项统计分析自闭症大脑重量与年龄相关变化的研究中发现,3-5岁男性自闭症患者的大脑重量比男性对照组高出15%(1451克对1259克)(图8)。成人大脑也有更多的脑脊液,这可能会错误地增加记录的大脑重量。即便如此,成人自闭症患者和非自闭症对照组大脑之间的差异也只减弱到1%左右(图8)。值得注意的是,3-5岁自闭症病例的平均脑重为1451克,与正常成年男性的平均脑重差不多(Courchesne et al., 1999)。
解剖学上的早期过度生长的区域:
正如下文所讨论的,迄今为止的研究表明,早期过度生长在整个大脑中并不是普遍存在的。到2-4岁时,一些区域和结构显示过度生长,而另一些则没有(Carper and Courchesne, 2005,Carper et al., 2002,Sparks et al., 2002)。虽然有许多可能的解释,但一种可能性是过度生长的相对时间或可能涉及的区域特定的遗传机制。
关于时间,可能有些区域在儿童早期或晚期没有显示过度生长的报道,因为这些区域的发育要早得多。也就是说,一些发育非常早的脑区实际上可能经历了异常加速的过度生长,但在过去MRI研究调查的年龄之前,即2-4岁之前。因此,有些区域或结构实际上可能在其他较晚发育的系统仍在经历过度生长的时间段内经历了随后的停止生长。鉴于此,有必要指出,异时性是所提出的阶段性假说中的一个关键变量。例如,枕叶皮层是一个早期发育的区域,正如接下来所讨论的,在对2岁以上的自闭症儿童的研究中,它是一个几乎没有过度生长证据的区域。目前,还没有关于2-4岁以下自闭症大脑的核磁共振研究,因此,例如枕叶皮层可能不会发生过度生长,或者过度生长可能在核磁共振研究考虑的年龄之前发生。
考虑到这些因素,对2-4岁自闭症儿童的MRI研究表明,额叶、颞叶和杏仁核是过度生长的高峰部位(见图6)(Carper and Courchesne, 2005,Carper et al., 2002,Courchesne et al., 2001,Hazlett et al., 2005,Sparks et al., 2002)。虽然整个大脑的灰质和白质体积大大增加(Courchesne et al., 2001,Hazlett et al., 2005),但在一些分别报告每个大脑叶灰质体积的研究和年龄中,一个引人注目的发现是,额叶和颞叶增大最多,枕叶最小(Bloss and Courchesne, 2007,Carper et al., 2002,Hazlett et al., 2006,Kates et al., 2004,Palmen et al., 2005)(见图9)。这种异常增大的区域梯度与认知功能受损最严重的区域相平行。在一项研究中,年龄较大的自闭症儿童的额叶和颞叶沟异常前移和/或上移(Levitt et al., 2003),这与额叶和颞叶的不成比例的增加是一致的。
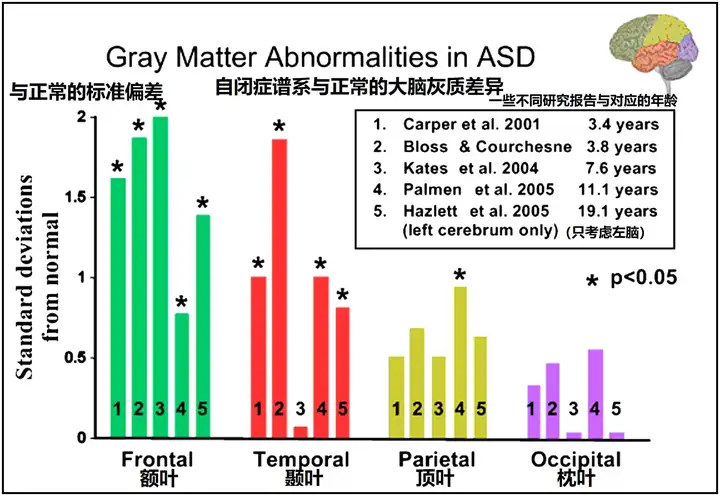
ASC大脑在早期以大大超过常人的规模发育,尤其是额叶(图9)。其中自闭症儿童前额叶皮质神经元数量达到了正常儿童的1.67倍(2至16岁的一些样本,参考Neuron Number and Size in Prefrontal Cortex of Children With Autism[11])。
前额叶被认为是大脑最高级的一些活动的生理基础:包括按照内部目标协调思想和行动,生存意志、个性的产生,执行功能(计划、决策、短期记忆、个性表达、调节社会行为和控制语言的某些方面,与区分相互冲突的想法、确定好与坏、更好与最好、相同与不同、当前活动的未来后果、为一个确定的目标而努力、预测结果、基于行动的期望以及社会“控制”等能力有关),更高层次的抽象的规则学习。
前额叶被认为是人类拥有超过其他物种智能的关键。文章Quantitative assessment of prefrontal cortex in humans relative to nonhuman primates[10]指出,人类的前额叶皮层占大脑的总量不成比例地大过其他物种。人类大脑皮层中前额叶所占比例是猕猴的1.9倍,是黑猩猩的1.2倍。白质中前额叶占的比例的差异甚至更加突出,人类是猕猴的2.4倍,是黑猩猩的1.7倍。ASC与正常人之间前额叶的差异已经达到了物种级别的显著性。在如此巨大的差异下,不合群和社交问题是可以想象的:与之类同的情况,想象独自一个人在黑猩猩群中出生并成长,抛开社交不谈,其大脑在这样的环境中能否正常塑造成人类的大脑,最终拥有正常人类的智能都是个问题,很可能成为了“傻子”。
这种前额叶皮层数量和体积上的优势能有多大程度转化为这些区域所负责的功能上的提升呢?抑或会因为“人类出生在黑猩猩群中”的原因而没有正常发展反而导致这部分功能紊乱呢?还是说这种大脑功能在量上的提升同时伴随着一些生理性的风险,这些风险导致了大脑功能无法正常发挥呢?一部分ASC战胜了环境和超级大脑带来的风险,掌控了自己的大脑,另一部分则在困惑与混乱中导致了该区域的功能发育受损,或者在两者之间反复横跳。他们一部分成为了历史上的超级天才(见第4章),一部分成为了典型自闭症,一部分有ADHD(注意力缺陷多动症),一部分患上了双相情感障碍,一部分出生自带抑郁症,一部分在天才和各种精神障碍之间反复横跳。作为一个非典型的大脑拥有者,在NT的世界中意识到这点并且用自己的方式掌控自己的大脑并非易事。本文第6章专门讨论这个问题,并研究如何战胜这些风险,掌握自己的大脑。
1.5 阿斯伯格和典型自闭症的神经学差异*
在DSM-V诊断标准中,阿斯伯格综合征和典型自闭症被归为更一般性的自闭症谱系状况。有争议的是它们虽然表现出了许多一致的特征(特别是高功能自闭症和阿斯伯格),但是否有着一些其他显著的神经学差异以至于需要一个额外的术语去研究呢?抑或者是阿斯伯格仅仅是一种程度较轻以至于没有语言发育迟缓的自闭症呢?之前我们都是讨论ASC一般性的神经学机理,这里我们讨论一下AS的特例。
文章Asperger's and Autism: Brain Differences Found报道了一项发表在BMC Medicine杂志上的研究,在脑电图检查中模糊地找到了阿斯伯格可能存在和自闭症的神经学差异:
研究研究员、波士顿儿童医院的神经学家Dr. Frank Duffy说:“阿斯伯格症儿童和自闭症儿童有相似之处:与发育正常的儿童相比,两者在大脑左半球一个被称为弓形筋膜的区域显示出较弱的连接,该区域涉及语言。”
然而,当研究人员观察大脑其他部分之间的连接时,他们看到了差异。阿斯伯格症儿童的左半球几个区域之间的连接性比自闭症儿童和正常儿童都要强。
Dr. Duffy说,阿斯伯格综合征儿童左半球脑区之间更强的连接性可能是阿斯伯格综合征患者在个性和能力方面的特殊之处。
耶鲁大学儿童精神病学教授Dr. James McPartland说:“这些发现令人振奋,方法也很复杂,他没有参与这项研究。虽然这项研究包括了合理数量的儿童,但像任何新发现一样,这项研究需要在未来的研究中进行复制。目前,很难知道新发现是否反映了阿斯伯格症和自闭症之间的核心、内在差异,或者是否反映了发展中的不同特征。”
AS左半脑某些区域有着比正常儿童更强的连接并非是一件令人震惊的事。因为在Saulnier & Klin, 2006[32]论文中对AS和HFA进行韦氏智商测试子分项对比时发现,尽管AS各言语和操作智力都高于HFA,但是言语理解部分智商分数显著地高得更多,比HFA高了22.5,甚至超过了正常儿童平均值15.6。这与对AS儿童的观察:非典型的学究式的语言使用方式吻合——也许AS儿童用某些特别的左半球大脑连接补偿了原本ASC语言的弱项。可能是阿斯伯格动用了他们高度发达的系统化能力(理性思考能力)来组织和理解语言,用系统化功能取代了大脑语言区的功能。
1.6 自闭症谱系的神经多样性
为了理解阿斯伯格,我们先来讨论阿斯伯格所属的一个大筐:自闭症谱系。
从严谨的科学角度来看,自闭症谱系并非由单一生物学特征引发的特定障碍,而是一大堆目前尚知之甚少的发育障碍的统称。这些障碍被归为一类,主要基于两个广泛的特征:社交障碍和刻板行为。这两种特征在程度上也存在很大差异,从非常轻微到严重差别很大。例如轻微的社交障碍可能就是表现比较特立独行不合群,而严重的可能行为逻辑完全超乎普通人能理解的范围。轻微的刻板行为可能只是爱好数学这类“系统化”的东西,严重的可能天天无法控制地拍手转圈圈。当然我们文章主要讨论的阿斯伯格是谱系最轻微的一类。
在谱系这个发育障碍大杂烩中,我们甚至尚未发现明确的共同的生物特征,仅仅有一些关于它们表征的统计趋势。目前已经发现了数百个与自闭症谱系相关的基因[34]。所以,今天可能会出来一篇论文发现某基因a与自闭症谱系风险相关,而明天又可能有人发一篇论文研究另一基因b如何影响自闭症谱系的发生。
由于涉及的变量众多且复杂,对这些障碍的分类也颇具挑战性。个体之间可能存在多个影响基因的组合,如abcde,或者相似但略有差异的基因组合,如abcdf。这导致了这些障碍之间看起来像是连续过渡的。因此,研究者只能根据这些障碍共同的社交不合群比较孤独的特征,将它们共同称为“孤独症谱系障碍”,并且解释说这是一种光谱一般丰富具有多样性的一类障碍(具体里面有啥不清楚,你就当它们像光谱一样啥都有,颜色之间还是连续的还没法分清楚就完事了)。因此“谱系”一词就诞生了。
也正因为目前谱系的如此宽泛的诊断对个体基本没有提供什么信息,对于谱系的所谓治疗本质只是对症治疗,如针对社交障碍进行社交训练,针对抑郁症治疗抑郁症状,针对一部分运动笨拙的改善运动能力等。什么,还有整天嫌老师讲课太简单上课大闹的?那你家长自己看着办,我也不知道咋办。
那有人就有疑问了,为何谱系这个大杂烩会有前面提到的“更敏感”“神经细胞长得更快”等特点呢?那是因为谱系虽然是个大锅,但就是根据刻板行为来筛选出来的,而轻微的刻板行为正是高智商的特征(包括理性、规则、擅长寻找规律等等)。可以预见谱系在神经生物学上会和高智商具有一些大致相同的发育方向。我们猜测那些智商不高的谱系可能是“物极必反”,刻板行为过于严重病态,并且一些帮助发挥大脑过度发育的辅助机制没有跟上,发育严重不平衡,反而阻碍了智力的发展。因此有学者认为这些严重的谱系可以视为高智力的发育失调[6]。从这个角度来说,和谱系相关的数百个基因可能都是某种“高智商预备基因”,它们的组合得好就会产生超高智商阿斯,而组合得运气不好可能就产生了典型自闭症。这部分内容详见第4章和第5章讨论。
剩下的筛选条件“社交障碍”是一个更加宽泛的条件,严格来说,任何与众不同的大脑理论上都会有不同程度的社交障碍,比如你和老鼠脑子差别太大,就没法社交,你和猩猩可能略微可以握个手击个掌社交一下。谱系属于大脑朝刻板(高智商预备)方向的与众不同,其与众不同的程度可能影响了社交障碍的程度。我们下一章就来讨论其中最为轻微的阿斯伯格是如何因此产生社交隔离感的。
2 阿斯伯格产生社交隔离感的原因
2.1 与众不同的大脑产生了换位思考的困难
先来复习一个心理学概念:
心智理论(Theory of Mind)是一种心理学和认知科学概念,指的是个体理解和预测他人思维、信念、情感和意图的能力。简单来说,心智理论就是我们理解和解释他人心理状态的能力,以便更好地预测和解释他们的行为。这种能力使我们能够在社交互动中表现出同理心和更好地与他人沟通。
我们可以把阿斯伯格社交困难的产生总结如下:
神经发育和常人不同—>思维方式和常人不同—>在同一个情境下的行为反应和常人不同—>如果把自己代入其他人的情境,假想自己做出的反应和其他人在这个情境实际做出的反应不同—>很难用换位思考的方法成功预测他人的行为(心智理论发展缓慢)—>只能通过观察并记忆别人在某种情况下的反应来预测他人的行为—>只能预测熟悉的人/已经见过的情况下别人的反应,只通过模仿已经学习到的别人的反应来使自己的行为像是普通人
阿斯伯格这种产生心智理论的方式和AI语言模型的训练很像,所以你会发现阿斯伯格和ChatGPT的心智理论、社交交流风格很相近。他们不是无法产生心理理论,而是以一种需要更加消耗脑力和更多数据量的方式产生心智理论。当然,这不代表他们社交能力会比普通人差,也许一些阿斯的脑力太强劲,又愿意花功夫去腾出一块算力训练心智理论,所以产生了比普通人更强的社交技能(比如一些顶尖政治家也是阿斯伯格)。虽然一些阿斯伯格的社交能力正常,但这种消耗,特殊的心智理论方式会产生一种社交距离感。这是因为普通人是通过“把对方当成自己换位思考”的方式社交的,而阿斯伯格们是通过“把对方当作一个不同于自己的物体”应用已学得的训练模型来进行社交的。如果没有事先了解阿斯伯格的知识,在周围人都是“亲切地”换位社交的情况下,就会感到一种不知道从何而来的莫名其妙的孤独感。
一些阿斯伯格之所以成为阿斯伯格是因为其智力远远强于普通人或者说大脑发育远远超前于普通人。Sheldon这种就是因为智力发育过于超常导致无法和普通人换位思考,这样从小长期在普通人之间成长起来,心智理论的发展就会非常缓慢,导致阿斯伯格。除了Sheldon这种天才导致的阿斯,还有一些智商普通的阿斯是由于“高智商预备基因”组合“歪”了导致脑发育“歪”了。这种“歪”只是一种不同的歪,并没有比正常人好或者差(就好像正常人的肚脐长在正面,但有些人长到反面去了),这种情况也会导致与众不同,从而导致社交障碍。如果说脑子发育得更加不同导致了一些社交之外的发展障碍,例如语言发育迟缓、认知发育迟缓、自理能力发育迟缓、(社交以外的)社会适应性发育迟缓,那就可能归为比较典型自闭症的范畴。
2.2 阿斯伯格的情绪超感知力
神经细胞的庞大复杂的丰富性导致了阿斯伯格的感官敏感。那么对于更加玄妙难以捉摸的情绪,AS的感知能力如何呢?会不会NT什么感觉都比AS迟钝但不知怎的唯独情感感知比起AS要敏感呢?随便想想都知道那当然是荒谬的。许多研究表明NT的情绪感知能力如同感官的感知能力一样,比起AS来说是非常迟钝的。
下面的一段译自Tony Attwood与Michelle Garnett所著的EXPLORING DEPRESSION AND BEATING THE BLUES - A CBT Self-Help Guide to Understanding and Coping with Depression in Asperger’s Syndrome [ASD-Level 1] [35]。
拥有 "第六感 "的情绪敏感性
阿斯伯格综合征的诊断特征之一是缺乏非语言交流:即阅读面部表情、身体语言和语气的能力。然而,临床经验和自传描述了一种感知和吸收他人负面情绪的“第六感”能力;此人对他人的痛苦、绝望、焦虑或愤怒过度敏感,而且这可能发生在他们没有真正看到或听到对方的情况下。一个例子是,一天早上,一个少年躺在床上,面对卧室的墙壁,闭着眼睛。他的母亲敲了敲门,默默地走进房间打开窗帘。他立即说:“怎么了,妈妈?”这是对她当时高度焦虑的情绪状态的准确评价,然而他并没有用常规的、非语言的线索来引出这一信息。
下面的引文描述了这种经历:
“对其他人的情绪状态有一种即时的、下意识的反应,多年来我对自己的情绪状态有了更深的了解。如果有人找我谈话,而他们充满了担心、恐惧或愤怒,我发现自己突然处于同样的情绪状态。”
“我能够分辨出别人看不到的非常微妙的线索,也可能是我从他们身上捡到的感觉。”
对他人强烈的负面情绪如此敏感,阿斯伯格综合征患者可能会被这种情绪“感染”,但却不知道自己为什么会有这种感觉。而且由于创造一种疏离感是如此困难,许多阿斯伯格综合征患者选择在社会上孤立自己,以保护自己的心理健康。
这种情绪敏感性也使得AS更容易焦虑抑郁(当然很大一部分原因归为被NT们排挤),Tony Attwood和Michelle Garnett提到:
阿斯伯格综合征患者似乎特别容易感到抑郁,大约有三分之一的青少年和三分之二的阿斯伯格综合征成年人在他们的生活中至少经历过一次严重的抑郁症。
许多阿斯伯格拥有非常强的同情心,能强烈地感受到他人的情感,但却由于与NT感受方式不同而无法理解NT为何出现了某种情绪,显示极弱的同理心。这种对他人社交感情直觉的不一致产生了心理理论的缺失。另外,情绪的高敏感也带来了情感管理的麻烦。Tony Attwood在《阿斯伯格完全指南》[36]中写到:
对阿斯伯格综合征人士来说,一个微小事件引起的愤怒情绪,其快速发作程度和强度往往非常强烈。
某些阿斯伯格综合征个体的情绪调节和控制机制似乎天生失灵,难以具备有效控制愤怒表达的能力。
这种情绪高敏感也使得多数适应社会良好的阿斯伯格都学会了切断情绪和意识行为的直接联系以防止行为失控过激,改用理性来做出在某种情况下“应当做出的”行为,例如什么时候要微笑、大笑、什么时候要做出严肃的表情都是理性分析的结果。但高敏感带来的情绪阈值易溢出的问题仍然能被观察到:人们经常描述AS个体只有两个情绪档位,即理性接管时的完全平静和超出理性控制时的激动暴怒。当AS处在完全放松的环境下(例如只有完全信任的父母在身边时),敏感的情绪才会不加修饰地表现出来。
2.3 情绪超感知力导致更快达到足够的社交情感需求
AS和NT的社交上的区别来源于AS的社交情感高敏感(或者也可说成NT社交情感的感知过于迟钝),如前面所说这可能源自神经联结更加丰富,以至于NT的日常社交对AS来说包含的情感过于刺激,会感到难受。这好比对光线敏感的人喜欢暗的环境,对声音敏感的人喜欢安静的地方,对社交情感敏感的AS更喜欢社交少、社交感情不强烈的环境,倾向于单独稳定可控的社交。但AS如同所有人类一样,保有基本的社交意愿和需求。AS需要社交,不过情绪超感知力导致AS比NT更快更容易地达到足够的社交情感需求,超过了需求后会感到厌烦。
在成长过程中社交感受与他人不同调,NT的社交情感过于强烈刺激,使得AS学会避开感情,来理性分析NT们的行为来解决和别人社交的问题。久而久之,外界NT们对情感的描述始终与自我感受无法对应,即各种感情词汇和情感预测理论都是描述NT的,和自己的感情不能相对应,AS自己的感情和学会的情感的语言描述产生了隔离,形成述情障碍。在接收的情感过多时有时会过载而发脾气,试图让周围在社交上“安静”下来。
AS的社交对NT来说缺少情感,是因为这样的交流已经产生了足够的情感感受了。另一方面这也使得AS的社交需求量大为降低,因为少量社交就可以满足社交情感需求。而NT需要不断大量的社交才可以满足其社交情感需求,这也使得NT把获得社交情感作为其活动的重要目的。而AS的活动就不会考虑过多的情感动机,只考虑有目的的实际理性效益,因此有时会非常不解NT的行为,觉得那些行为没有意义,那都是没考虑到大量的社交才能满足NT的社交情感需要。最终行为动机导向了NT会花更多功夫研究社交,发展各种各样的社交方法,把社交做得更加复杂刺激,而AS对社交没有花那么多功夫,预测NT的理性分析理论也越来越跟不上NT的花里胡哨社交的发展。
2.4 用吃作一个形象的例子
用吃的做例子,AS就像对单位食物吸收能量的能力更强的人,和普通人一起吃东西,吃一点就吃饱不吃了,并且普通人吃的全是糖的甜甜圈给AS吃实在太重口,吃着难受。另一方面,AS也需要吃东西获得足够的能量,可是普通人发现每次约AS出来吃东西他吃一点就停下不吃要赶紧走人,也不能理解普通人一把一把吃炸鸡的乐趣,自顾自吃蔬菜沙拉,和一般人完全不搭调,之后他们就不约AS一起吃了。结果好巧不巧在这个虚构的世界中“吃饭”必须和别人一起才能吃,于是找不到一起吃饭的人的AS虽然饭量小,反而被饿死了。
为了和其他人一起吃饭补充能量不饿死,AS必须学着装作和大家一样大把大把抓炸鸡吃,然后塞进嘴不咽下去,不去想这个炸鸡的味道,因为这味道太重口了(屏蔽抑制住自己的社交感情),再找机会偷偷吐出来,只从桌上拿套餐里的蔬菜沙拉吃。AS经过学习人云亦云也会和大家一起讨论起炸鸡的美味了,知道用什么词来形容吃炸鸡的感觉(用理性分析社交),大家也都约AS出来吃了,但有时候换家店AS还大搬之前那家店那一套说辞,说这个炸鸡真辣,但明明新的店的炸鸡是甜的,众人笑AS吃不出味道。随着人们对炸鸡鉴赏能力的提高到处山吃海吃,AS再也装不出炸鸡鉴赏家的样子了,但AS却找到了一个肯天天陪他吃米其林的NT朋友,在一家熟悉的店天天品鉴着一小碟一小碟精致的菜,每盘味道都控制得很精确微妙。虽然这个NT朋友也尝不出这些菜之间的细微区别,觉得都尝不出味道,但这个朋友知道AS是好人,也崇拜AS的一些特异功能,就陪着他一起一顿又一顿吃着。当然,这个朋友和AS吃饭总是吃不饱,他还有其他一些NT朋友可以一起吃饭,所以主要是靠其他朋友吃饱的。但他对于AS却是唯一的可以一起吃饭的人,没了他AS就会饿死。
有时候有NT和AS之间小打小骂,那个NT直接抓起一把做的难吃的料理朝AS嘴里塞,但他没想到AS尝起来那个料理就等价于粪便,以为只是一般难吃开开玩笑。AS直接呕吐出了来恶心了好几天,并且用超级记忆力一直记得那天被喂了翔。
2.5 另一个例子:四色视觉
世界上约有1%的女性是拥有四种视锥细胞,她们在光谱识别上多了一个橙色频率的识别点。因此她们有红绿蓝橙四原色而非普通人的红绿蓝三原色,比普通人能识别出更多的颜色差别。但普通人的世界所有屏幕和印刷都是基于三原色的,所以她们会因为现实看到的颜色和人们描述的以及屏幕书本显示的颜色不同而苦恼。她们有着更精确的颜色分辨能力,对于她们来说红绿混搭的黄和纯黄在她们看来是两种颜色。同时一些普通人看来平淡的场景在她们看来是颜色鲜艳过于刺激的。(参考BBC的科普The women with superhuman vision)
这跟AS的情况有一些相像。AS体会到的感情和普通人的情感的不一样,AS的对感情的分辨力更敏感,所以只能生硬地记住别人所说的情感是对应自己的哪种。就像四色视觉的人要把黄色和(红+绿)都强行记成黄色,要理性地记住,而非直接感到这两个都是黄色。成长的路上还很疑惑别人为什么把颜色那么分类,完全没有道理。就好像我们三色视觉的人生活在一群只看到黑白的人当中,看着他们把浅蓝和深红都叫做深灰一样奇怪。这种方法的结果就是AS们只能把自己的情绪用NT的语言简单地归类为“还行”和“糟糕”,不知道如何用学到的NT的语言来描述自己的感受,结果就是产生了述情障碍。那些没有经过训练的四色视觉的人久而久之也会把自己感到的一切都塞到三原色的框架里描述了,有许多不太明显的四色视觉者一辈子也没发现自己是四色视觉的,只是学会按照外界语言来描述一切颜色。