图说英国新冠变异株
凡人发了一篇很全面的介绍[英国的新冠突变]的博文,给大家提供了交流互动的好机会。 言尤末尽,想到一图至少顶10句 (A picture is worth a thousand words), 就来图说一下这个之前称B.1.1.7 现被命名为 'VUI-202012/01'的英国新冠变异株吧。
十七个氨基酸变异位点的分布:
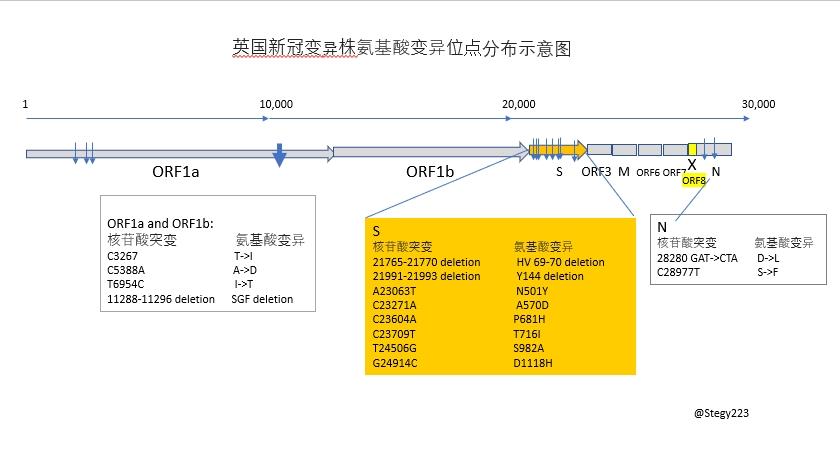
决定每个氨基酸突变对蛋白质分子结构与功能的影响最主要有二点:
1) 突变发生在蛋白质分子三维结构的那个位点?
2) 什么性质的突变?要谈这点就得了解一下生物化学。20种氨基酸从每种的分子结构和化学反应性上讲大体可分为5大组:
- 疏水性不与其它氨基酸分子作用的
- 亲水性易与其它氨基酸分子形成氢键的
- 带正电荷的,帶负电荷的
- 还有Pro 是唯一侧链为环状的。

蛋白质分子比较能接受同类型的氨基酸变异(Conservative substitution) 。发生在蛋白质三维结构不同位点的不同类型的变异就有的可接受有的不太可接受。上述二点结合起来有些氨基酸突变对蛋白质分子的结构和功能可以没什么影响, 有些可以多多少少有一定程度影响,个别影响巨大。
众知周知新冠刺突S蛋白是决定入侵细胞的关键蛋白,最终表现为传染性的强弱。mRNA疫苗是基于对S蛋白与宿主细胞识别与结合的机理而量体裁衣设计的。所以目前关心的重点是S蛋白上的突变。就用个S蛋白三维结构示意图来探讨一下这一新冠变异株中S蛋白上的8个突变。

S蛋白在病毒膜外有三大块。NTD亚基的功能不是太清楚,相比S1和S2亚基,进化上不是很保守,序列中突变多一些。S1亚基负责与宿主细胞膜上ACE2受体结合,S2亚基负责完成与细胞膜融合从而将病毒基因组RNA弄进宿主细胞。
2个删除突变(HV69-70 deletion 和 Y144 deletion)发生在NTD亚基上,应该不会影响到争对S1亚基产生的抗体。
S1亚基受体结合区域N501Y突变最受关注。之前在小白鼠动物模型实验上已发现N501Y突变增强传染性。N501是直接与ACE 受体结合的关键氨基酸之一。所以可以比较肯定地推测这一变异株的N501Y会具有同样的效果。
仅N501Y突变应该不会太影响争对S1亚基产生的多克隆抗体,因为这些不同的抗体分子各自识别S1亚基的不同部位(
另外受到密切关注的是P681H突变。P681H在S1与S2亚基交接部位,紧邻弗林酶(Furin)切位点PRRARS -> HRRARS。尚难判断有多大影响。可能影响不大。
S蛋白之外,最值得一提的是发生在ORF8 上的一个转译终止符突变 (Q27Stop)。ORF8 是新冠编码的病毒特异的辅助蛋白之一。新冠病毒辅助蛋白有ORF3, ORF6, ORF7和ORF8。己知ORF8对病毒复制非必需但有协助病毒适应宿主的功能。好象欧洲大陆不怎么闹蝙蝠和冠状病毒。
毫无疑问各国的科学家们正在开足马力从各个层次(
更多我的博客文章>>>