5月里经朋友介绍才注意到文学城中这场关于葡萄糖转运机理“发明权”及关于颜宁在Nature2012或Nature2014 (1,2) 中是否需要引用润涛阎Cell1993/PNAS1995(3, 4)的大讨论(5, 6, 7)。自5月以来烽火连三月,这一争论俨然演变成了一个科学公案。近来讨论的焦点逐渐集中到了Nature2014作者在讨论部分提出的(proposed) working model中的葡萄糖载体蛋白构象循环图(图五)。此处作者简单提及“according to the Alternating Access Model ”. 一篇相关文献即Jardetzky Nature1966出现在前言中。
从Jardetzky66直接进入2014文中图5提出的四象循环图确实留下了让人提问的空间。小颜微博中的对老先生66年的经典你们要怎么着式的回答自然不光没有帮助解答问题反而更使人感到此处有点迷雾重重了。
从科学的角度细看看这66经典和构象循环图究竟是什么关系是个很有意思的话题。网友SwipeTheFox 在这方面做了有益的努力(8). 有趣的是在与Fox 先生的讨论中一不小心俺觉得看出了这里面一些规律,好象在文献中还没出现过另外也没人谈及。俺就也顺便提出(propose)一个MFS 转运蛋白构象转换模型(Model) 吧。
1. MFS大家庭转运蛋白构象
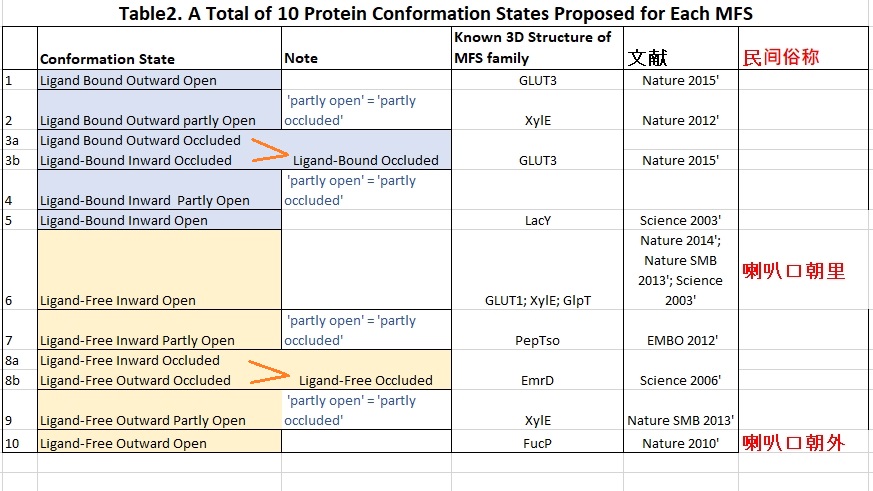
综观有关的文献MFS转运大家庭转运蛋白构象可用/(至少)3个参数来描述,即底物/糖结合与否,喇叭口开关状态? 喇叭口朝外还是朝里(细胞内) (Table1):
a. 底物/糖结合与否只有二种可能: Ligand-bound, ligand-free。
b. 喇叭口朝外还是朝里自然只有二种可能。
c. 喇叭口开关状态需至少3个值即全开,全关或半开半闭。当然“半开半闭”仍是个粗略的描述,完全可能有其它中间状态。
这样 2x2x3 共有至少12种可能的构象状态。考虑到当喇叭口全关闭时无所谓朝里朝外,所以10种构象应是比较全面的描述了(Table2).
2. Alternating Access Model (AAM).
老先生在1966 到底说了啥? 老先生从对动力学数据的分析提出底物结合位点应当是朝外开和朝内开轮流着来 (AAM)。当时的认知完全到不了涉及多少个转运蛋白构象的程度。关于AAM 与构象转换模型之间的关系小颜甚至YGS 大师分别在2013年的综述里(9, 10)详细讨论了. 阅读学习后受益非浅,这里不赘述。否则如果只是简单地抱着Jardetzky1966那不成了原教旨主义者了。
3. MFS 10构象转换模型
这10个构象自然不是随便画在一起就可以的。这些构象之间必须一步步变化过来,因为转运蛋白是镶嵌在双磷脂细胞膜海洋里,蛋白整体可以在膜里打转转,蛋白本身构象也可高速转换(thermodynamics) 但绝不能来个后空翻朝细胞外与朝细胞内来回翻跟头。所以在符合AAM 基本思想的情况下上述10构象应当遵守下面提出的构象转换模型(Figure1):

需要提出的是这一新的10构象转换模型自然涵盖9构象,8构象,7构象 ...
4. 关于构象转换动力学研究
获得人葡萄糖载体高精度3维结构的成果标志着对于人葡萄糖载体转运机理的研穷从此走出了“瞎子摸象”的阶段。从多个构象的对比研究中确能在分子水平上获得许多重要的insight (11). 同时对转运机理的进一步深入研究仍需要Biochemical, biophysical, kinetic 及蛋白结构等各种实验手段的协同作战。对“活的”蛋白的动力学研究(老阎多次强调的)是个重要课题。“活的”载体蛋白在无底物/糖存在的情况应在下列至少3个构象之间变换。那变换的速率有多快?在达到平衡状态下这“活的”载体蛋白喜欢处于那一种构家呢? 等等的问题(Figure2) 都有待于新时代里更多的biochemical, biophysical, kinetic 实验来问答
业余人士搞科研,疏漏之处在所难免。敬请业内专家学者批评指正。如果这一模型根本性地错了,俺一定撒掉拙文。如果本模型经检验是站得住脚的,希望这一模型有助于对糖转运机理更进一步的研究。
参考文献:
1. Sun, L., et al “Crystal structure of a bacterial homologue of glucose transporters GLUT1-4” Nature 2012. 490:361-366
2. Deng, D., et al “Crystal structure of the human glucose transporter GLUT1” Nature 2014. 510:121-125
3. Yan, R.T. and Maloney P.C “Identification of a residue in the translocation pathway of a membrane carrier” Cell 1993. 75:37-44
4. Yan, R.T. and Maloney P.C “Residues in the pathway through a membrane transporter” PNAS 1995. 92:5973-5976
5. 润涛阎: 颜博士的回应 https://blog.wenxuecity.com/myblog/1666/201905/17824.html
颜宁的反驳为何站不住脚 https://blog.wenxuecity.com/myblog/1666/201905/19103.html
颜宁博士无法反驳的第五个事实 https://blog.wenxuecity.com/myblog/1666/201907/7515.html
6. Fanreninus: 阎欠颜一个道歉 https://www.wenxuecity.com/blog/201907/73963/7931.html
7. SwipeTheFox: 阎颜之争风云再起 http://blog.wenxuecity.com/myblog/49113/201907/9857.html
8. SwiperTheFox: 转运蛋白一日游 http://blog.wenxuecity.com/myblog/49113/201907/17275.html
9. Yan, N “Structural advances for the major facilitator superfamily (MFS) transporters” Trends in Biochemical Sciences 2013, 38:151-159
10. Shi, YG “Common folds and transport mechanisms of secondary active transporters” Annu. Rev. Biophys. 2013. 42:51-72.
Note: This paper covers MFS famility. Section ‘Alternating Access Model’ starts from Page 58.
11. Deng, D., et all “Molecular basis of ligand recognition and transporter by glucose transporters” Nature 2015. 526:391-396
更多我的博客文章>>>