摘自《尼安德特人》
在波恩莱茵博物馆的尼安德特人上的一小块肩胛骨。多年来我们得到过太多令人失望的结果,所以我并不抱太大希望。无论我们怎样提取,得到的十之八九都是自其出土约 140 年来渗入到骨头中的细菌或人类的 DNA。但在电话里,马蒂亚斯听起来很激动。他真的提取到了尼安德特人身上的遗传物质?还是别抱过多期望为好。
来到实验室,我发现马蒂亚斯和拉尔夫·施米茨(Ralf Schmitz)在一起。这位年轻的考古学家曾帮助我们从存放在波恩的尼安德特人化石中取到一小块肩胛骨。当他们给我看一串从测序仪中得出的 A、C、G、T 序列时,这两人都情不自禁地笑了。我和他们以前都不曾见过这样的序列。
对于外行来说,这似乎只是一个由四个字母组成的随机序列,事实上,它们是 DNA 化学结构的简明表示,而作为遗传物质的 DNA 几乎存在于身体的每个细胞。DNA 的双螺旋结构为人们所熟知,其中的两股链由核苷酸腺嘌呤、胸腺嘧啶、鸟嘌呤和胞嘧啶组成,分别缩写为 A、T、G 和 C。这些核苷酸的排列顺序储存着让我们身体成形并维持各项功能运作的遗传信息。我们所研究的特殊 DNA 片段是线粒体基因,即 mtDNA,它经由母亲的卵细胞转递给后代。线粒体 DNA 的数百份拷贝都储存在细胞内的微小结构——线粒体中,并且这些 DNA 携带的特定信息对于线粒体的产能来说十分必要。我们每个人都只携带一种线粒体 DNA,它只占了我们基因组的 0.0005%。由于我们的每个细胞均携带着成千上万个同类型线粒体 DNA 的拷贝,所以特别容易研究。它不像我们携带的其他的 DNA 只有两份拷贝,一份来自母亲一份来自父亲,且均存储在细胞核内。截至 1996 年,我们已研究了几千份来自世界各地的人类线粒体 DNA 序列。通常这些序列会被拿来与第一个已确定的人类线粒体 DNA 序列进行比较,因而这个常见的参考序列可以用来编译列表,展示不同位置的具体差异。让我们大喜过望的是,从尼安德特人骨中得到的序列所包含的变化,不曾出现在之前研究过的数千份人类 DNA 序列中。我简直不敢相信这是真的。
每每得到激动人心或意想不到的结果时,我的心中便会充满怀疑。我会仔细检查所有出错的可能。也许有人用牛皮制成的胶处理了骨头的某个部位,所以我们才会在实验结果中发现牛的线粒体 DNA。但是这种可能很快被否定了。我们立即检查了牛的线粒体 DNA(已经由别人完成测序),发现两者之间存在非常大的差异。这个新的线粒体 DNA 序列显然非常接近人类的序列,但与已经测过的几千份人类 DNA 序列相比还是略有不同。我开始相信,这确实是首个提取并测序自一种已灭绝人类的 DNA 片段。
我们打开一瓶存放在实验室咖啡厅冰箱里的香槟。我们知道,如果我们所看到的真的是尼安德特人的 DNA,那么自此便开启了无限可能。也许有一天,我们真的能比较尼安德特人和现存人类的所有基因或任何特定的基因。当我穿过漆黑静谧的慕尼黑走回家时(我喝了太多香槟无法开车),我简直不敢相信所发生的事情。回到床上,我辗转反侧无法入睡。我一直在想尼安德特人的事情,以及我们刚刚获得的那个线粒体 DNA 样本。
图 1.1 重建的尼安德特人骨架(左)以及现代的人类骨架(右)。照片来源:肯·幕布雷(Ken Mowbray),布莱恩·梅利(Blaine Maley),伊恩·塔特索尔(Ian Tattersall),加里·索耶(Gary Sawyer),美国自然历史博物馆。
1856 年,达尔文的《物种起源》出版的前三年,在杜塞尔多夫以东约 10 千米处的尼安德特河谷,工人在清理采石场的一个小山洞时,发现了一个头盖骨和一些骨头。他们认为这些骨头来自熊。但几年之后,这些遗骸被鉴定为来自一种已灭绝的人类。这是首次有人描述此类遗骸。此发现震惊了博物学界。多年来,关于这些骨头的研究一直在持续开展,并且发现了更多类似的骨头。这些研究想要知道尼安德特人是谁?他们是如何生活的?他们为什么在大约 3 万年前消失?在欧洲和尼安德特人共存的数千年间,我们现代的祖先与他们是如何互动的,他们是朋友还是敌人?尼安德特人是我们的祖先,抑或我们失联已久的表亲(见图 1.1)?尼安德特人行为特征方面的迷人细节对我们而言并不陌生,如照料伤患、举行葬礼仪式,甚至创作音乐等。考古遗址的发掘结果告诉我们,相较任何现今的猿类,尼安德特人与我们更相像。那么,到底有多像呢?他们是否会说话?他们是否是人类家族演化分支中走入末路的一个物种?抑或是,他们的一些基因流传至今,现在仍隐藏在我们体内?这些问题都已成为古人类学的重要课题。可以说这个学科领域在那些骨头从尼安德谷发现之时便开始建立,而现在已经可以从那些骨头中得到遗传信息了。
这些问题本身就足够有趣。不过在我看来,尼安德特人的骨头片段会带来更大的惊喜。尼安德特人是现代人类最为近缘的已灭绝的亲属。如果研究他们的 DNA,我们无疑会发现他们的基因和我们的非常相似。几年前,我的团队对黑猩猩基因组中的大量 DNA 片段进行了测序。结果表明,在我们人类与黑猩猩共有的 DNA 序列中,只有略高于 1% 的核苷酸存在差异。显然,尼安德特人肯定比这个结果更接近于我们人类。但是(这令人倍感欢欣鼓舞),我们在尼安德特人的基因组中找到的这些差异,其中一定有一些会将我们区别于早期的人类祖先。这些祖先不仅仅是尼安德特人,还有生活在大约 160 万年前的图尔卡纳男孩(Turkana Boy)、大约 320 万年前的露西(Lucy)以及 50 多万年前的北京人(Peking Man)。也正是由这些少数差异构成的生物学基础,使现代人类诞生之后又演化出了全新的行为模式,包括出现迅速发展的技术、我们如今所熟悉的艺术形式,以及目前已知的语言和文化。如果可以研究尼安德特人的 DNA,那么我们便可以解开以上所有谜题。怀揣着这样的梦想(或幻想),我终于在旭日东升时进入梦乡。
第二天,马蒂亚斯和我都较晚才到实验室。检查完昨晚的 DNA 序列,确保我们没有犯任何错误之后,我们坐下来,计划下一步该做什么。从尼安德特人化石中得到一小段看起来有趣的线粒体 DNA 序列是一回事,但要让我们自己信服这是一个生活在(在如此特殊情况下)大约 4 万年前的人类的线粒体 DNA,又完全是另一回事,更不用说让世界上的其他人都相信了。过去 12 年的工作经验让我清楚地知道下一步该如何做。首先,我们必须重复试验——不只是最后一步,而是所有的步骤,从提取一块新骨头开始,从而证明我们所获得的序列并非来自骨头中严重损坏和历经变化的现代线粒体 DNA 分子。其次,我们必须延伸线粒体 DNA 序列,这些序列是通过骨头提取物的重叠 DNA 片段而得到的。这样我们能够重建一个更长的线粒体 DNA 序列,从而开始估计尼安德特人的线粒体 DNA 与当今人类相比是多么不同。接下来的第三个步骤也是必需的。我自己经常要求,来自古老骨骼的 DNA 序列需要经由特别的证据证实——即在另一个实验室重复试验。在竞争尤为激烈的科学领域,这是一个不同寻常的步骤。我们肯定会因为宣称获得了尼安德特人 DNA 而被视为异类。为了排除实验室中未知的错误来源,我们需要与一个独立的实验室分享一些珍贵的骨头材料,并希望他们能重复我们的结果。我与马蒂亚斯和拉尔夫讨论了所有想法。我们制订了工作计划,并彼此发誓在研究团队之外,每个人对于这项研究绝对保密。在确定我们所获的结果真实无误之前,我们不想引起关注。
马蒂亚斯立即开始工作。他曾花了近三年时间试图从埃及木乃伊中提取 DNA,不过均徒劳无果。这次前景看好,他信心满满。拉尔夫回到波恩后似乎有些沮丧,因为他只能在那里焦急地等待我们的结果。我试着专注于手头的其他项目,但我很难将马蒂亚斯在做的事完全抛于脑后。
马蒂亚斯要做的事情并非都那么容易。毕竟,我们处理的不是从活人血液样本中提取的完整而纯净的 DNA。教科书中干净利落的双链螺旋 DNA 分子,其核苷酸 A、T、G、C 以两股糖-磷酸骨架互补配对(腺嘌呤与胸腺嘧啶,鸟嘌呤和胞嘧啶)。当储存在细胞核和细胞线粒体之中时,DNA 不是一个静态的化学结构,相反,DNA 不断受到化学损伤、被复杂的机制识别和修复。此外,DNA 分子非常长。细胞核中的 23 对染色体中的每一个都包含一个巨大的 DNA 分子。一组 23 条染色体全部加起来大约有 32 亿对核苷酸。由于细胞核有两份基因组拷贝(每份拷贝存储着一组 23 条染色体,分别继承自我们的母亲和父亲),所以细胞核中包含约 64 亿个核苷酸对。相较之下,线粒体 DNA 太小,只包含约 16500 个核苷酸对。但考虑到我们的线粒体 DNA 是古老的,因而测序的挑战极大。
无论是核 DNA 还是线粒体 DNA,最常见的自发损伤都是胞嘧啶核苷酸(C)失去氨基,然后变为一个核苷酸;这个核苷酸不是 DNA 自然产生的,它被称作尿嘧啶(简称为 U)。细胞中有酶系统。酶系统会去除这些 U 并替换成正确的 C。丢弃的 U 最终成为细胞垃圾。通过分析随尿液排出的受损核苷酸,我们计算出每天每个细胞大约有 1 万个 C 变成 U,这些都要被移除并加以更换。这只是我们基因组遭受的几种化学攻击之一。例如,核苷酸会丢失,产生空的位点并导致 DNA 分子链迅速断裂。在断裂发生之前,有些酶会填补丢失的核苷酸。如果发生断裂,其他酶会将 DNA 分子重新结合在一起。事实上,如果这些修复系统不复存在,我们细胞中的基因组连保持 1 个小时的完整状态都做不到。
当然,这些修复系统的运作需要能量供给。我们死后会停止呼吸,体内的细胞耗尽氧气,也就无法制造能量。DNA 的修复一旦停止,各种损伤会迅速积累。除了活细胞中不断发生的自发化学损伤,一旦细胞开始分解,很多死亡后的损伤也会开始出现。活细胞的重要功能之一是保持酶和其他物质相互分离隔断。有些隔断中含可以切割 DNA 链的酶,这些酶对于某些类型的修复而言十分必要。其他隔断中含有可以分隔 DNA 与各种微生物的酶,这些微生物有的会进入细胞,有的会被细胞吞入。一旦生物体死亡并耗尽能量,隔断膜就会恶化分解,这些酶就会泄漏,并开始不受控制地降解 DNA。死亡后的几小时到几天内,我们体内的 DNA 链被切割成越来越小的碎片,而其他各种形式的损伤也逐步累积。同时,当我们的身体无法维持原有的隔离细菌的屏障时,生活在我们肠道和肺部的细菌开始失控生长。这些过程将一起最终摧毁储存在我们 DNA 中的遗传信息——这些信息曾调控我们身体的形成、持续运转以及各项功能。历经完这个过程,我们便失去了彰显生物独特性的最后一道痕迹。从某种意义上说,我们的肉体已彻底死亡。
不过,我们身体中的几万亿个细胞,每个都几乎包含整套 DNA。因此,只要身体的某个角落有一些细胞内的 DNA 逃过被完全分解的过程,那么就会留存下遗传痕迹。例如,酶降解和改变 DNA 的过程需要水才能运作。如果我们身体的某些部分在 DNA 降解之前就变干燥,酶降解和改变 DNA 的过程就会停止,我们的 DNA 片段有可能会保存很长时间。这种情况是可能发生的。例如,躯体存放在干燥的地方而变成了木乃伊。这样的全身干燥有时是意外发生的,取决于生命终结时所处的环境;也可能是刻意为之。众所周知,古埃及人经常将死者做成木乃伊。在大约 5000 至 1500 年前,为了能给他们的灵魂提供死后的栖身之所,数十万人的尸体被做成了木乃伊。
即使没有变成木乃伊,身体的某些部位,如骨骼和牙齿,可能在尸体埋藏之后长期保存。这些硬组织含有细胞,这些细胞位于用显微镜才能看到的小孔中,负责骨折后新骨的生成。当这些骨细胞死亡,它们的 DNA 可能外渗,并与骨头的矿物成分结合在一起,从而阻止酶的进一步攻击。因此,幸运的话,有些 DNA 可以避开身体死亡后的降解和损伤侵袭而残存下去。
但是,即使有些 DNA 在死亡后的身体乱战中幸存下来,其他进程仍会继续降解我们的遗传信息,尽管速度较慢。例如,来自太空的背景辐射不断冲击地球,进而产生了修改和破坏 DNA 的活性分子。此外,一些过程需要水的参与才能进行(如 C 失去氨基,生成 U),即便 DNA 被保存在相对干燥的条件下,这些过程仍将持续。因为 DNA 有亲水性,因此即使在干燥的环境中,水分子还是会附于两股 DNA 链之间的沟槽中,让需水的自发性化学反应得以发生。C 失去氨基(去氨基)是其中最快的过程,它会破坏 DNA 的稳定,并最终打破 DNA 链。大部分这样或那样的过程,仍然未知。它们会不断瓦解在细胞死亡浩劫中幸存下来的 DNA。虽然破坏速率取决于许多因素,诸如温度、酸度等。但很清楚的是,即使在最好的条件下,使人之为人的遗传程式的残存信息终将被摧毁。那块经过我和同事分析的尼安德特人骨头,即便已经历 4 万年之久,所有这些过程还未完成终极破坏任务。
马蒂亚斯得到了一个序列长度为 61 个核苷酸的线粒体 DNA。要做到这一点,他必须得到此 DNA 片段的多个拷贝。在这个过程中,他用到了聚合酶链反应(PCR)。为了证实我们的发现,他从重复初次所做的 PCR 实验着手。这个实验要用到两条很短的合成 DNA 片段,我们称之为引物(primer)。设计引物的目的是结合线粒体 DNA 的两个部位,并让 61 对核苷酸分开。这些引物与从骨骼中提取的少量 DNA 以及 DNA 聚合酶混合,这种酶能以引物为起点和终点,合成新的 DNA 链。加热这个混合物使两条 DNA 链解链,然后在混合物降温之时,A 与 T 配对、G 与 C 配对,引物便能与目标序列结合。酶会以引物与 DNA 链结合为起点,合成 2 股新链、复制骨头中原有的 2 股链,这样 2 股原始链便变成了 4 股。这样不断重复扩增,可制造出 8 股、16 股、32 股等,总共可重复三四十次。
美妙绝伦的 PCR 技术威力巨大,由特立独行的科学家凯利·穆利斯(Kary Mullis)于 1983 年发明。原则上一个 DNA 片段经过 40 个周期之后可获得约万亿份拷贝,这才使我们的研究成为可能。所以在我看来,穆利斯理应获得诺贝尔化学奖,而 1993 年他的确实至名归地得到了。然而,PCR 的高灵敏度也使我们的工作变得困难。从一个古老的骨头中获得提取物,其中可能含有极少数幸存的古 DNA 分子,或者根本就没有,甚或是包含一个或多个现代人的 DNA 分子,而这些 DNA 分子会污染实验:它们可能来自我们使用的化学品、实验室的塑料制品或空气中的灰尘。在人类生活或工作的房间里,大多数尘埃颗粒中都含有人体的皮肤碎屑,而皮肤碎屑中都是满含 DNA 的细胞。另外,处理骨头的人,如博物馆的工作人员或挖掘人员,他们的 DNA 也可能污染样品。正是基于这些方面的考虑,我们选择研究尼安德特人线粒体 DNA 中差异最多区域的序列。由于许多人的序列在这个特定的区域有所不同,我们至少可以知晓有几个人的 DNA 纳入了我们的实验之中,并觉察出其中的差错。这就是为什么我们看到一个前所未有的 DNA 序列变化会如此兴奋。如果序列看起来与当今人类相似,我们无法确定这到底意味着尼安德特人与当今人类的线粒体 DNA 确实是相同的,抑或是我们找到的只是隐伏于某处(如一粒尘埃)的当今人类的线粒体 DNA 片段。
我对污染这事太了解了。我从事古 DNA 的提取和分析方面的工作超过了 12 年。古 DNA 主要来源于那些已灭绝的哺乳动物,如洞熊、猛犸象以及大地懒等。得到一连串令人沮丧的结果(在所有用 PCR 分析的动物骨头中,几乎都检测到了人类线粒体 DNA)之后,我花了很多时间思考和设计方法,把污染降到最小。因此,马蒂亚斯在一个保持得尤为干净,且与实验室其余部分完全分离的小实验室里进行所有的提取和其他试验,直到 PCR 的首个温度循环。把古 DNA、引物以及其他必要的 PCR 组分都一齐放入试管中,并将试管密封起来,然后再将温度循环和随后的所有实验移至常规实验室进行。在洁净实验室里,每周都用漂白剂冲洗一次所有东西的表面;每天晚上都用紫外线照射实验室,以破坏尘埃中携带的任何 DNA。马蒂亚斯进入洁净实验室之前必须通过一个前厅,他和其他人在那里穿上防护服、防护面罩、发网和无菌手套。所有试剂和仪器都被直接送到洁净实验室,研究所其他地方的任何东西都不允许带入实验室。马蒂亚斯和他的同事每天在洁净实验室开始一天的工作,而不是在我们实验室的其他地方(那里正在分析大量的 DNA)。一旦进入实验室的其他地方,这些人当天就不许再进入洁净实验室。说得婉转一点,我对控制 DNA 污染已近乎偏执,且觉得理应如此。
即便如此,在马蒂亚斯一开始的实验中,我们还是看到了一些受现代人类污染的证据。在使用 PCR 扩增骨头中的线粒体 DNA 片段之后,他在细菌中克隆出了一批 DNA 拷贝——它们理应完全相同。他这样做是为了观察克隆出的分子中是否含有多种线粒体 DNA 序列。每个细菌中一段长达 61 个核苷酸分子的序列都会与一种名为质粒的载体分子结合,然后克隆出数百万个细菌。每个克隆均携带首个细菌包含的 61 个核苷酸分子的拷贝。所以通过测序大量克隆,我们能够概览分子群体中存在的 DNA 序列差异。在马蒂亚斯最初的实验中,我们看到了 17 个彼此相似或相同的克隆分子,它们与已知的 2000 多个现代人类线粒体 DNA 有所不同(我们加入现代人类的线粒体 DNA 进行比较)。但我们也看到其中一个序列与某个当今人类的序列相同,这清楚地表明污染的存在。污染也许来自博物馆馆员或骨头发现至今 140 多年来曾处理过它的其他人。
所以,为了重现原始结果,马蒂亚斯首先便要重复 PCR 和克隆。这一次,他发现了 10 个带有独特序列的克隆,这让我们兴奋不已。还有 2 个克隆应该来自现代人类。然后他用骨头再提取了一次,也做了 PCR 和克隆,得到了 10 个有趣的克隆以及 4 个看似是现代人类的线粒体 DNA。现在我们很满意:我们的原始结果已经通过了第一项测试。我们可以重复结果,每次重复都能看到同样独特的 DNA 序列。
接下来,马蒂亚斯开始“沿着”线粒体 DNA 一鼓作气,用设计好的其他引物扩增与第一个片段有部分重叠的片段,而这个片段会进一步延伸到线粒体 DNA 的其他区域(见图 1.2)。我们再一次从这些片段的部分序列中观察到从未在当代人类中出现过的核苷酸改变。在接下去的几个月里,马蒂亚斯扩增了 13 个大小不同的 DNA 片段,每个片段至少重复两次。要想解释这些序列谈何容易,任何一个 DNA 分子会因为各种各样的原因而携带突变:曾经的化学改变,测序错误,或者仅仅只是某个人的某个细胞中的线粒体 DNA 分子出现了罕见的自然突变。因此,我们使用了我先前研究古动物 DNA 时所采用的策略(见图 1.2)。我们能在每次实验的每个位置上找到确切的共有核苷酸——就我们检测的大部分分子而言,该位置上总携带有特定的核苷酸(A、T、G 或 C)。我们也要求在两个独立实验中,每一个位置都是相同的。这是因为在极端的情况下,PCR 可能只从一条单一的 DNA 链开始复制。在这种情况下,由于首次 PCR 循环中的一些错误,或特定 DNA 链的一些化学改变,所有的克隆会在同一位置带有相同的核苷酸。如果在一个位置出现两次 PCR 差异,我们就再重复做第三次 PCR,观察哪种核苷酸会再次出现。马蒂亚斯最终用 123 个克隆的 DNA 分子拼凑出线粒体 DNA 变异最大的由 379 个核苷酸组成的序列。根据我们之前已经确定的标准,这便是这个尼安德特人生前携带的 DNA 序列。一旦有了这个较长的序列,我们就可以开始激动人心的工作:将它与现代人类存在的变异进行比较。
图 1.2 尼安德谷的尼安德特人的线粒体 DNA 片段重构。第一行是现代人类的参考序列,下面的每一行代表从尼安德特人标本扩增而来的一个克隆分子。在这些序列与参考序列相同的地方,我用点来标示;在它们的核苷酸存在不同的地方,我就写出来了。底部那行是重建的尼安德特人核苷酸序列。每个位置与参考序列的不同,我们均要求至少在两个独立的 PCR 实验中(至少显示其一)的大多数克隆中看到。Matthias Krings et al., “Neandertal DNA sequences and the origin of modern humans,”Cell 90, 19–30 (1997)。
此时,我们将包含 379 个核苷酸的尼安德特人线粒体 DNA 序列与来自世界各地的 2051 个现代人的线粒体 DNA 序列进行比对。尼安德特人和现代人类之间平均存在 28 个不同位点,而现代人类彼此之间平均只存在 7 个差异。尼安德特人线粒体 DNA 与现代人类的差异是现代人类之间的差异的 4 倍。
接下来,我们想要寻找一切可表明尼安德特人线粒体 DNA 与现代欧洲人线粒体 DNA 更为相像的迹象。有人可能会很想找到这样的证据,毕竟尼安德特人曾在欧洲和亚洲西部演化和定居。事实上,一些古生物学家认为,尼安德特人是当今欧洲人的祖先之一。我们将尼安德特人的线粒体 DNA 与 510 个欧洲人的线粒体 DNA 进行比较,发现平均存在 28 个差异。接着我们将尼安德特人的线粒体 DNA 与 478 个非洲人以及 494 个亚洲人的线粒体 DNA 进行比较,他们的线粒体 DNA 平均差异也为 28 个。这意味着,以平均差异而言,欧洲人的线粒体并不比现代的非洲人和亚洲人的更接近尼安德特人。但也有人认为,尼安德特人遗传了一些线粒体 DNA 给某些欧洲人,因此这些欧洲人中的线粒体 DNA 可能更接近尼安德特人的线粒体 DNA。我们进行了检查并发现,样本中最像尼安德特人的欧洲人线粒体 DNA,存在 23 个差异;最接近尼安德特人的非洲人和亚洲人分别存在 22 个和 23 个差异。总之,我们发现尼安德特人的线粒体 DNA 不仅非常不同于全世界现代人类的线粒体 DNA,且没有任何迹象表明,尼安德特人的线粒体 DNA 和现今某个欧洲族群的线粒体 DNA 有着任何特殊的关联。
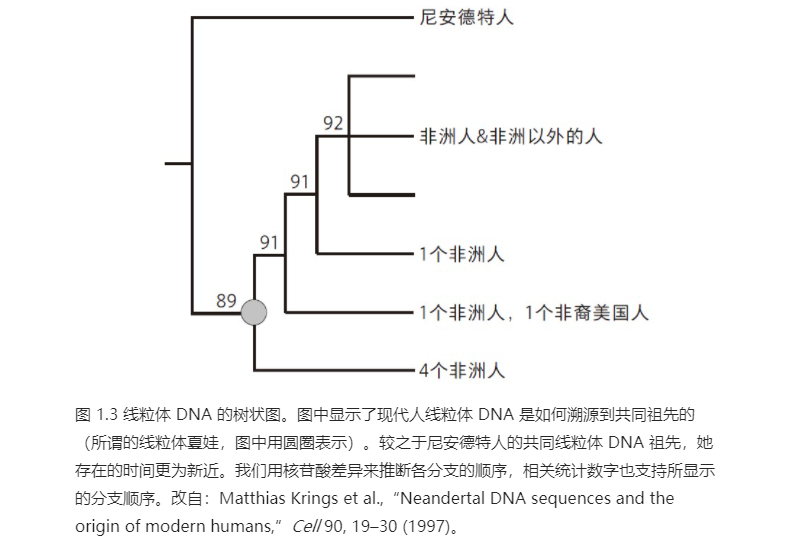
然而,仅依据计算出的差异数目还不足以重建一段 DNA 的演化历史。DNA 序列之间的差异代表了过去发生的突变。不过某些突变会更频繁地发生,并且 DNA 序列中的某些位置也更易发生突变。在 DNA 序列的演化史中,这些位置可能发生过不止一次突变,尤其针对那些更为频繁发生的突变。因此,为了评估这个特殊线粒体 DNA 片段的历史,我们需要用模型来模拟线粒体 DNA 如何变异和演化,并谨记某些位置可能发生过不止一次突变,因此会掩盖先前的突变。这番重建的结果可以用一张树状图来呈现。分支顶端的 DNA 序列是共同祖先的 DNA 序列,树上侧分支的连接点为古 DNA 序列(见图 1.3)。当重建完这样一张树状图时,我们发现,现今所有人类的线粒体均可溯源到一个共同的线粒体祖先。

图 1.3 线粒体 DNA 的树状图。图中显示了现代人线粒体 DNA 是如何溯源到共同祖先的(所谓的线粒体夏娃,图中用圆圈表示)。较之于尼安德特人的共同线粒体 DNA 祖先,她存在的时间更为新近。我们用核苷酸差异来推断各分支的顺序,相关统计数字也支持所显示的分支顺序。改自:Matthias Krings et al.,“Neandertal DNA sequences and the origin of modern humans,”Cell 90, 19–30 (1997)。
早在 20 世纪 80 年代,这一发现就因艾伦·威尔逊(Allan Wilson)的研究而为人所知。1对于线粒体 DNA 而言,这一追溯结果早在预料之中。因为我们每个人都只有一种线粒体 DNA,且无法与群体中的其他人交换线粒体 DNA 分子。线粒体 DNA 只能通过母亲传递,如果一位女性没有女儿,她的线粒体 DNA 血脉就会断绝,因此每一代中都有一些线粒体 DNA 血脉消亡。这也意味着,曾经一定有这么一个女人(所谓的线粒体夏娃),她携带线粒体 DNA 血脉,是当今所有人类线粒体 DNA 的共同祖先——但这纯属偶然,因为其他所有血脉自那时起都因为各种原因断绝了。
但是,根据我们的模型,尼安德特人的线粒体 DNA 没有回溯到这位线粒体夏娃,而是回溯到更早以前的现代人类的共同祖先。这一发现令人欣喜过望。毫无疑问,这表明我们的确已经找到尼安德特人的 DNA 片段;同时还指出,至少在线粒体 DNA 层面,尼安德特人和我们大相径庭。
我和同事一起用该模型估计尼安德特人线粒体 DNA 与现代人类线粒体在多久之前开始拥有共同祖先。这两种线粒体 DNA 的差异数目表明了它们之间的代际时间长短。相隔很远的物种的突变速率(如小鼠和猴子)会有所不同,但非常相近的物种之间(如人类、尼安德特人和类人猿)的突变速率很稳定。根据看到的差异,科学家足以估计出两份 DNA 序列最晚拥有共同祖先的时间。通过线粒体 DNA 中不同类型突变的速率模型,我们估计目前所有人类的线粒体 DNA 祖先(线粒体夏娃),生活在 20 万至 10 万年前,这恰好与艾伦·威尔逊及其团队的发现相吻合。然而,尼安德特人线粒体 DNA 和现代人类线粒体 DNA 的共同祖先则生活在大约 50 万年前,也就是说,比现今人类线粒体 DNA 的祖先(线粒体夏娃)还要古老 3~4 倍。
这个发现很棒。我现在完全相信,我们已经得到了尼安德特人的 DNA,它与现代人类的 DNA 非常不同。然而,在公布这项发现之前,我们需要克服最后一道障碍:我们需要找到一个独立的实验室,重复我们所做的事情。这个实验室不需要确定所有 379 个核苷酸的线粒体 DNA 序列,只需要得到一个突变区域,这个区域携带有使尼安德特人与当今人类区别开来的一个或多个突变。这样才能证明,我们确定的 DNA 序列真的存在于骨骼之中,而不是漂浮在我们实验室里的一些奇怪和未知的序列。但是,我们能找谁帮忙呢?这是个微妙的问题。
毫无疑问,许多实验室都想参与这样一个颇具前景且吸引人眼球的项目。不过我们有风险:如果我们挑选的实验室不像我们那样努力减少污染和解决其他所有有关古 DNA 的问题,实验人员可能无法成功提取并扩增到相关序列。如果发生了这样的事,我们的结果会被认为是不可重复的,因而无法发表。我知道没有人能像我们这样,花大量的时间和精力在这类工作上,不过我们最终还是选择了美国宾夕法尼亚州立大学群体遗传学家马克·斯托金(Mark Stoneking)的实验室。马克曾在伯克利的艾伦·威尔逊实验室攻读研究生并做过博士后。我在 20 世纪 80 年代后期做博士后时就结识了他。他是发现线粒体夏娃的幕后功臣之一,也是现代人类起源的“走出非洲”学说的构建者之一。“走出非洲”学说认为现代人类于 20 万至 10 万年前起源于非洲,然后分散到世界各地,没有杂交便取代了所有早期人类,如欧洲的尼安德特人。我敬重他的判断和正直,也知道他是一个随和的人。此外,他的一个研究生安妮·斯通(Anne Stone),曾于 1992~1993 年间在我们实验室工作。安妮是一位认真且雄心勃勃的科学家,曾与我们一起从美国原住民的遗骸中获取线粒体 DNA,所以熟知我们的技术。我觉得如果有人能重复我们的结果,非她莫属。
 选择“Disable on www.wenxuecity.com”
选择“Disable on www.wenxuecity.com”
 选择“don't run on pages on this domain”
选择“don't run on pages on this domain”